Oculomotor Unit Diversity | Neural Control of a Complex Oculomotor Plant (NIH/NEI, PI: JM Miller)
Motor Unit Diversity in Horizontal Eye Movement Control (NIH/NEI, PI: PD Gamlin)
In it’s first 40 years, oculomotor physiology did quite well without a method for measuring eye muscle forces, and In place of such measurements made assumptions, a central one being the oculomotor final common path hypothesis (FCP), which supposed a fixed relationship between ensemble motoneuron (MN) firing rate and muscle force (at a given muscle length) irrespective of type of eye movement (Robinson 1968, 1975). Given FCP, however, the finding that abducens MN activity was higher in converged gaze (for a given position of eye in orbit; Mays & Porter, 1984; Gamlin et al, 1989; Zhou & King, 1998) implied co-contraction of lateral rectus (LR) and medial rectus (MR) muscles.
Does co-contraction actually occur? We developed chronically-implantable muscle force transducers (MFTs) to measure physiologic eye muscle force, and showed that, on the contrary, LR and MR muscle forces were slightly lower in convergence (Miller et al. 2002). This was called the missing force paradox because the increased force expected with generally increased MN firing was not found. Miller, Davison & Gamlin (2011) brought the issue to a head by simultaneously measuring both muscle forces and unit activity in the abducens nucleus and the MR subdivision of the oculomotor nucleus. Consistent with earlier findings, they found that abducens neurons decreased activity less during convergent than during conjugate adduction (mean kv/kc = 0.50, where kv is the slope of the rate-position curve for vergence, and kc for conjugate movement at distance), which implies stronger LR innervation in convergence. Further, they found that MR MNs increased activity more during convergent than during conjugate adduction (mean kv/kc = 1.53), also implying stronger MR innervation in convergence. Despite stronger innervation in convergence at a given eye position, they found both LR and MR muscle forces to be slightly lower in convergence, -0.40 g and -0.20 g, respectively. Evidently, the relationship of ensemble MN activity to total oculorotary muscle force depends, so to speak, on the position of the other eye.
With FCP disproved, it is no longer possible to assume that MNs compose a homogeneous “pool”, varying only in threshold and sensitivity, similarly recruited by every eye movement subsystem. We believe instead that “paradoxical MNs” (LR MNs with kv < kc and MR MNs with kv > kc) innervate muscle fibers that are weak, have mechanical coupling that attenuates their effective oculorotary force, or serve some non-oculorotary function. That is, oculomotor motor units (MUs; each being an MN and the muscle fibers it innervates) have diverse functions. It is already clear that some MUs don’t directly rotate the eye (eg, orbital fibers couple to the tendon only indirectly; Oh, 2001), and that nonlinear interactions among muscle fibers (eg, Goldberg et al, 1997) are probably the norm. Given the specialized characteristics (contractile speed, fatiguability, laminar location) of the 6 or so conventionally distinguished EOM fiber types, and that all fibers of an MU are apparently the same type (Porter et al. 1995), it would make functional sense for different types of units to be recruited by different eye movement subsystems.
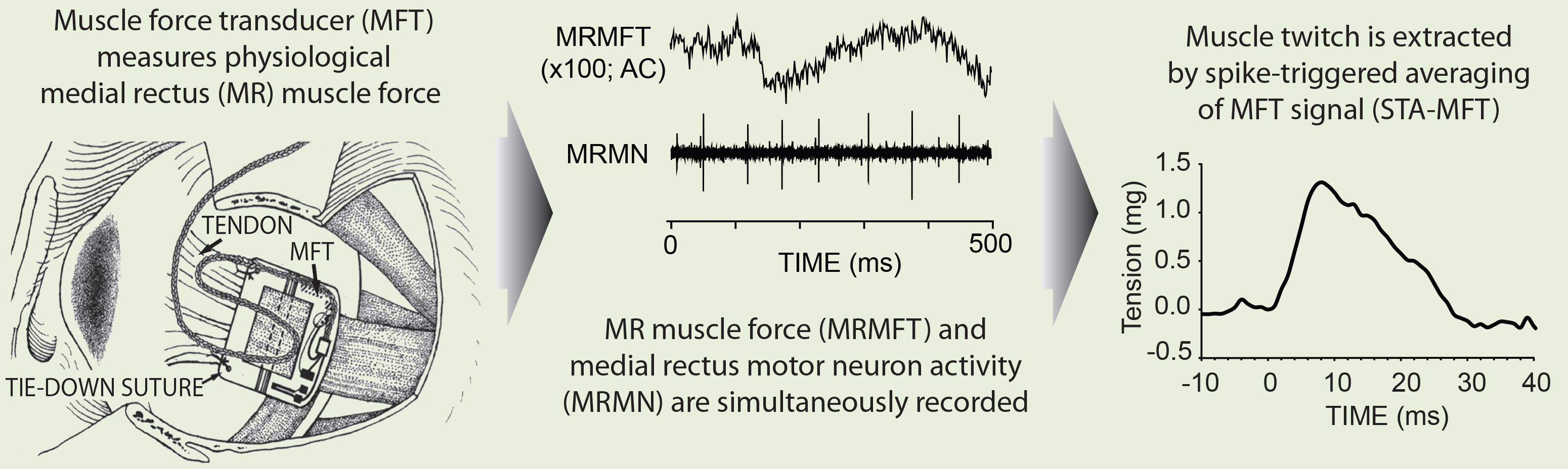
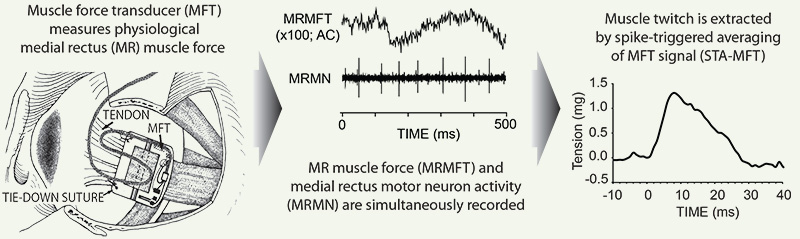
We are therefore keen to characterize (oculorotary force, speed, fatiguability) individual MUs and then determine their participation in various types of eye movement. We have taken the first steps, using MN recordings to perform spike-triggered averaging on muscle force signals (STA-MFT), and demonstrating that during steady fixation with low firing rates it is possible to extract individual MU twitches. We demonstrated the STA-MFT technique in recordings from 33 medial rectus MNs, from which we were able to identify and measure the twitch characteristics of 20 MUs (Gamlin & Miller 2012). It would then possible, as in conventional studies, to determine an MN’s activity during eye movements at higher firing rates, but now with knowledge of the MU’s oculorotary mechanical characteristics.
Drugs that modify eye muscle properties, elsewhere under development, show promise as simple, inexpensive office procedures able to effect corrections not possible with strabismus surgery. Our STA-MFT technique would be suitable to study the fiber type specific functionality of pharmacologically-modified motor units in alert animals
Gamlin PD, Miller JM (2012). Extraocular muscle motor units characterized by spike-triggered averaging in alert monkey. Journal of Neuroscience Methods, vol 204, pgs 159-167 (Authors' Cut PDF; Publisher's site).
Miller JM, Davison RC, Gamlin PD (2011). Motor nucleus activity fails to predict extraocular muscle forces in ocular convergence. Journal of Neurophysiology, vol 105, pgs 2863-2873 (Paper PDF).
Miller JM, Bockisch CJ, Pavlovski DS (2002). Missing Lateral Rectus Force and Absence of Medial Rectus Co-Contraction in Ocular Convergence. Journal of Neurophysiology, vol 87, pgs 2421-2433 (Paper PDF). |
|